

2022年10月26日,Nature Communications在线刊出了北京师范大学认知神经科学与学习国家重点实验室邢大军课题组的题为“V1-bypassing suppression leads to direction-specific microsaccade modulation in visual coding and perception”的论文。该研究通过记录猕猴初级视皮层(V1)和V2的跨层活动结合行为学实验,首次发现猕猴在注视过程中,微眼跳的产生对V2的神经活动及视知觉敏感性的动态调制具有方向特异性。
Background
日常生活中,人和其它灵长类动物通过快速转移视线来高效地探索周遭环境。即使当目光长久聚焦在一个点上时,我们的眼睛依然会以每秒2~4次的频率产生突然的微小运动。这种不自主的小幅运动即为“微眼跳”。微眼跳虽小到难以被主观察觉,却能在感知上引起强烈的视觉效果,比如引发运动错觉(图1a)、刷新视觉信息以及抵抗视觉影像的消退(图1b)。
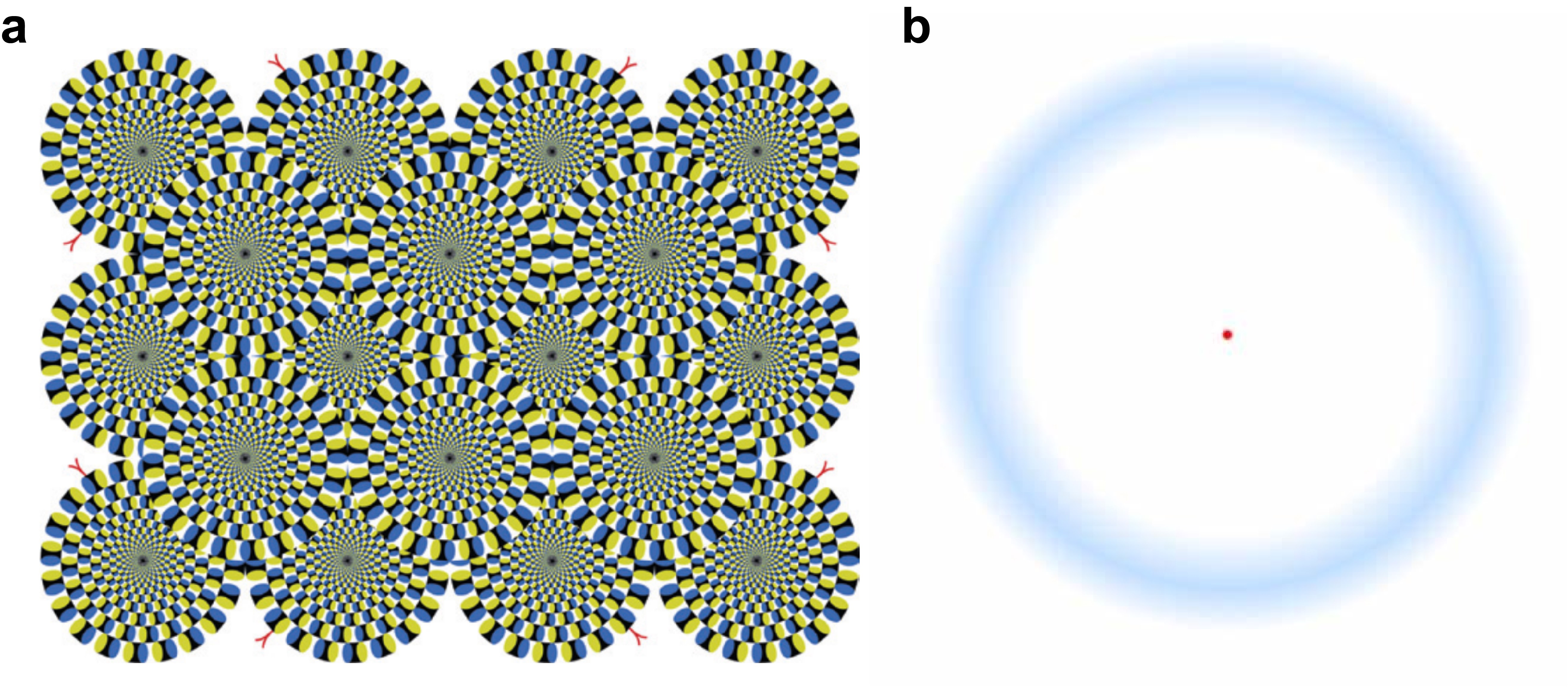
图1: a. 运动错觉。盯住圆环的中央黑点,依然会感受到圆环的运动,这是因为微眼跳的存在,每发生一次微眼跳就会引发运动错觉1。b. 视觉消退错觉(Troxler’s effect)。盯住中央红点5-10秒,会发现外周蓝色光晕逐渐消失,移动眼睛之后又重新出现。事实上每产生一次微眼跳,消退的影像也会短暂地复现2, 3。
正如《达芬奇密码》中所言:“The mind sees what it chooses to see.”眼睛只看向大脑选择的事物4, 5。那么微眼跳是否也在我们的潜意识中,默默地对视觉信息进行着选择呢?近年,来自人的行为学研究对于微眼跳方向是否调制视知觉存在矛盾性的发现,有研究者认为各个方向微眼跳所产生的效果没有差别3,也有研究者发现微眼跳方向和视知觉表现存在联系6, 7。现象级争议只是冰山一角,背后隐藏的神经机制仍迫切等待研究和回答。
Results
微眼跳的方向如何影响视觉皮层信息加工及其与行为的关系是当前研究的首要问题。对此,作者记录了V1、V2在猕猴微眼跳产生前后的神经活动(图2a-b),首次揭示出微眼跳与神经元感受野的相对方向对V2群体反应的特异性影响(图2c中蓝线):朝向V2神经元感受野的微眼跳引发的放电反应强于远离方向的微眼跳所引起的反应,并且在连续的相对方向变化过程中呈现出梯度变化。
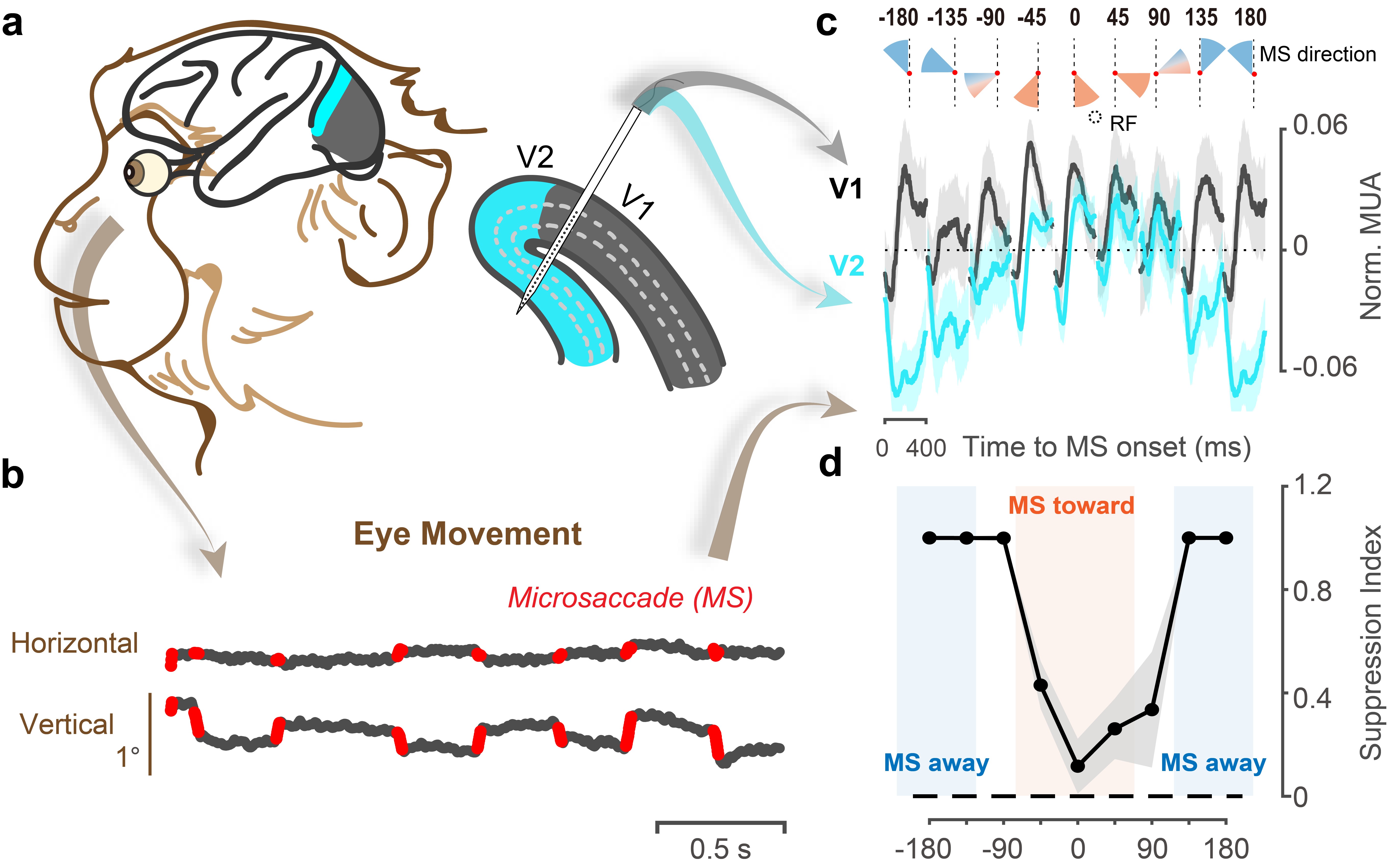
图2:a. 线性电极记录V1以及其下方覆盖的V2脑区示意图。b. 从眼动轨迹中提取出微眼跳。c. 不同微眼跳方向对应的V1、V2群体放电活动。d. 不同微眼跳方向对应的V2抑制性强度。
与V1中微眼跳引起的各向同质的反应模式相比(图2c中黑线),V2受到的方向特异性调制主要表现为远离感受野微眼跳带来的抑制作用(图2d)。此外,这一抑制作用的强度和微眼跳的幅度存在显著相关,暗示着V2的方向特异性抑制很可能源于脑内动眼系统的运动信号。
为了进一步探究这种方向特异性调制的环路来源,作者提出了两种来源假设:一是来自前额叶部署眼动相关脑区的反馈性投射;二是来自控制微眼跳的上丘核团的皮下通路投射。前者的投射连接坐落在V2表层而后者位于V2中间层。利用这一解剖学分层差异,作者通过描绘V1、V2不同层在微眼跳发生前后的时空动态反应(图3a),结果发现V2中间层最早出现方向性调制(图3b),且其调制强度也最高,支持了方向特异性抑制的皮下起源(图3c)。
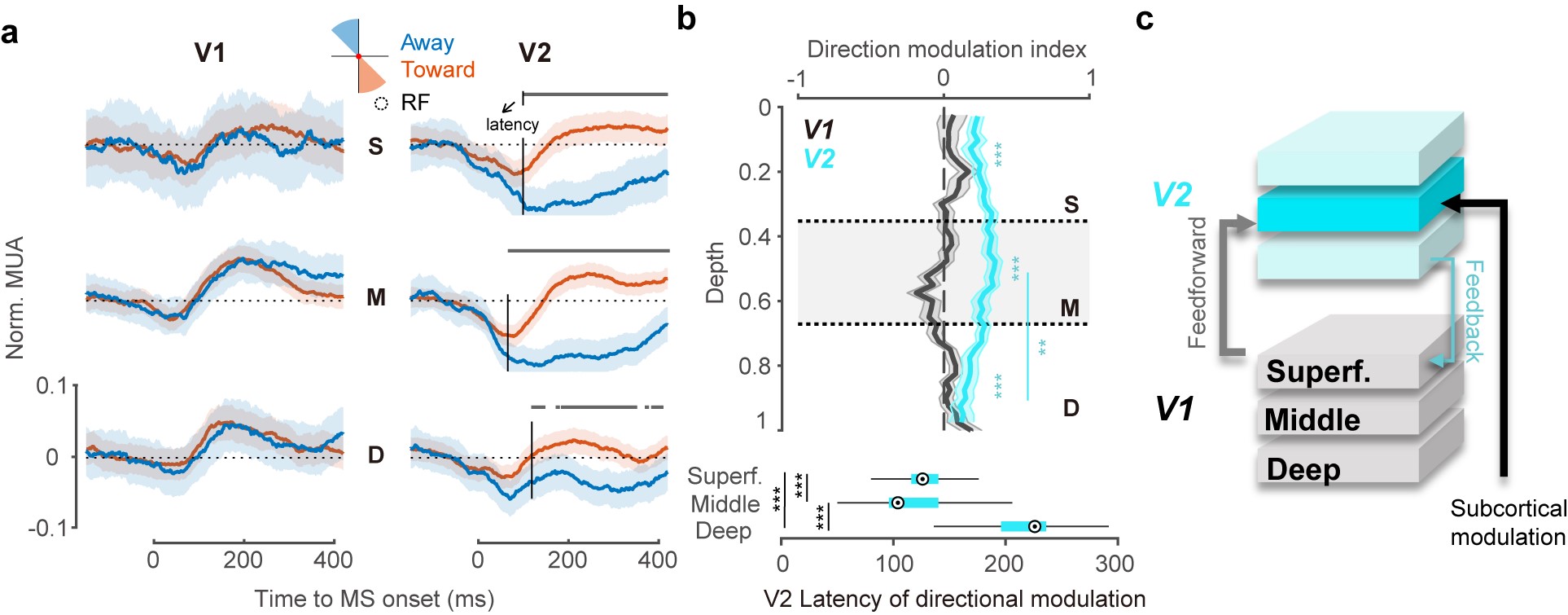
图3:a. V1、V2的表层、中层和深层对两类方向微眼跳的反应。b. 方向性调制强度跨层变化及其出现时间的跨层分布。c. 环路机制示意图。
上述发现意味着,一个向左看的“小动作”会使得左脑的V2脑区活动被抑制(左脑视皮层接收右视野信息),令其变得不太“兴奋”了。那么,左右脑区不平衡的兴奋状态是否让左右视野的视觉敏感性也出现差异呢?于是,作者设计了一个目标检测任务,训练两只猕猴对一个快速闪现且非常微弱的小亮斑进行检测并报告出亮斑的位置(左视野or右视野),同时记录下猕猴的微眼动和行为选择。有趣的是,如果在目标亮斑出现之前的0.3-0.7秒内,猕猴产生了一个朝向目标方向的微眼跳,相比于远离目标方向的微眼跳,猕猴对目标的检测正确率更高。这说明视觉敏感性的动态变化确实依赖于微眼跳的方向。
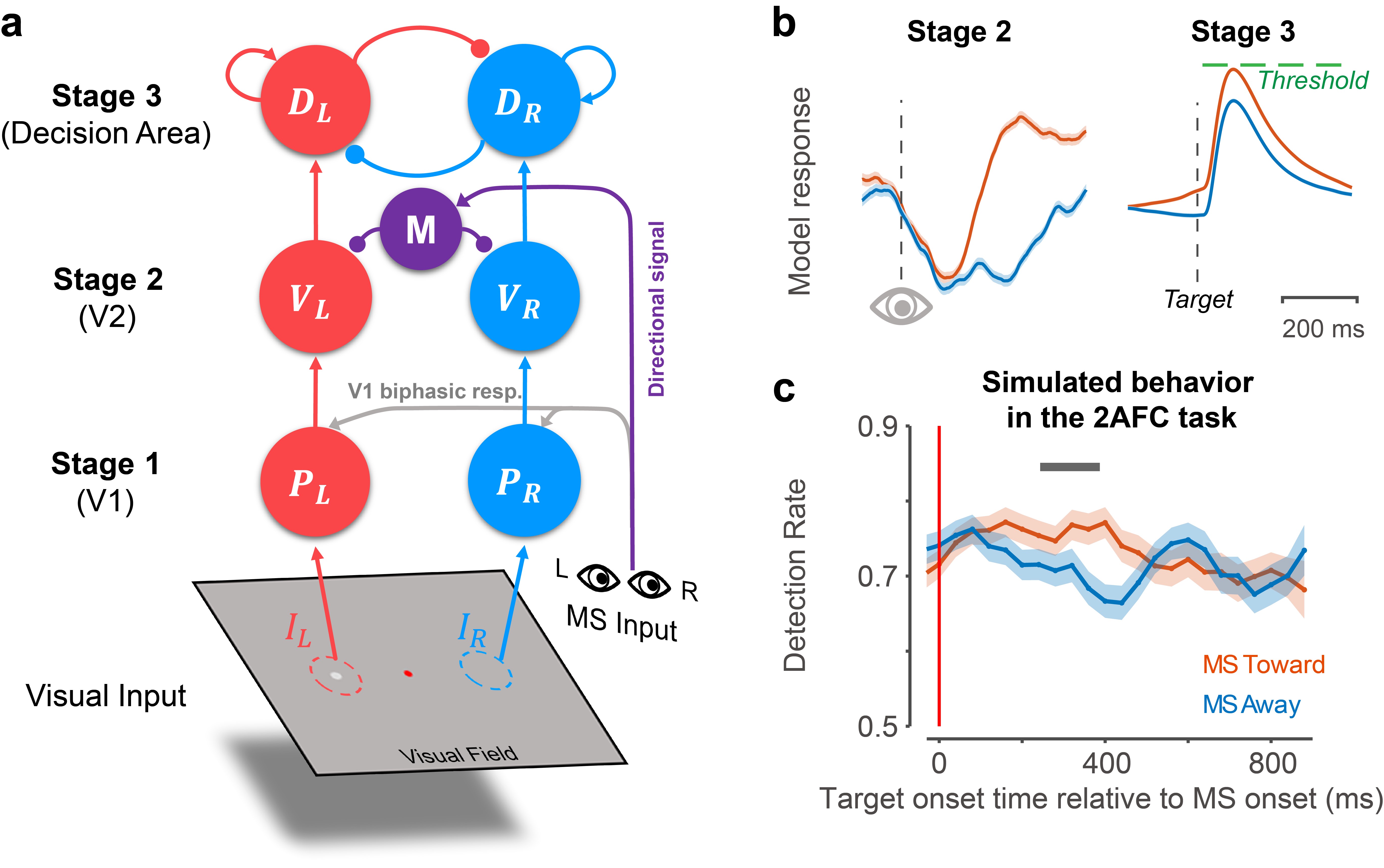
图4:a. 动态决策模型示意图。b. 左侧为模型第二阶段对朝向感受野和远离感受野方向的微眼跳的反应。右侧为第三阶段朝向和远离微眼跳发生后对目标亮斑的反应。c. 仿真目标检测任务中的行为动态
在论文的最后一部分,作者依据电生理发现建立了动态决策模型(图4),为微眼跳方向性调制视觉敏感性提供模型解释。在引入了微眼跳的方向性抑制信号(图中紫色成分M)后,模型不仅复现出了V2的反应模式(图4b),还仿真出了猕猴产生微眼跳之后的行为动态变化及方向差异(图4c),并解释了两只猕猴的个体差异。动态决策的三阶段模型将当前研究的电生理发现和行为结果联系到了一起,进一步说明了微眼跳的方向特异性抑制机制的作用。
Summary
综上,这项研究发现了猕猴微眼跳对V2的方向特异性调制,并揭示出该机制引起的视觉感知能力的时空动态变化。微眼跳带来的方向性调制可能源于皮下通路所传递的眼动相关的运动信号,这一运动信号以抑制的形式,首先作用在V2的中间层中感受野位于微眼跳相反方向的神经元群体,进而借助前馈连接继续影响到下游视觉系统。一次向左的微眼跳发生后,V2的兴奋性在左右视野空间上出现差异,使得左右脑视觉系统对新输入进来的弱信号具有不同的传递效力,右视野的弱信号更容易在传递过程中逐渐衰减,更难到达意识层面,最终导致了左右两侧视觉敏感性的变化。
这项工作挑战了人们对微眼跳的固有认识,发现在不需要空间注意的主动参与时,大脑对视空间的信息选择就已经存在了。该文作者强调了微眼跳虽然在尺度上仅局限于中央视野1度范围,但对外周视野的感知和决策也具有深远的影响。这对广大基于注视的认知实验具有直接的参考意义。
该研究得到了国家自然科学基金(32171033,32100831)、中国博士后科学基金(2021M690435)、北京师范大学博一交叉学科基金(BNUXKJC1909)、中央高校基本科研基金、111项目基金(BP0719032)以及认知神经科学与学习国家重点实验室开放课题基金的资助。该论文第一作者为博士生武宇洁,通讯作者为邢大军教授。课题组博士后王天及其他成员对此项工作也做出了重要贡献。
全文链接?:https://www.nature.com/articles/s41467-022-34057-3#Sec9
1. Otero-Millan, J., Macknik, S.L. & Martinez-Conde, S. Microsaccades and blinks trigger illusory rotation in the "rotating snakes" illusion. J Neurosci 32, 6043-6051 (2012).
2. Martinez-Conde, S., Macknik, S.L., Troncoso, X.G. & Dyar, T.A. Microsaccades counteract visual fading during fixation. Neuron 49, 297-305 (2006).
3. McCamy, M.B. et al. Microsaccadic efficacy and contribution to foveal and peripheral vision. J Neurosci 32, 9194-9204 (2012).
4. Engbert, R. & Kliegl, R. Microsaccades uncover the orientation of covert attention. Vision Res. 43, 1035-1045 (2003).
5. Hafed, Z.M. & Clark, J.J. Microsaccades as an overt measure of covert attention shifts. Vision Res. 42, 2533-2545 (2002).
6. Shelchkova, N. & Poletti, M. Modulations of foveal vision associated with microsaccade preparation. Proc Natl Acad Sci U S A 117, 11178-11183 (2020).
7. Bellet, J., Chen, C.Y. & Hafed, Z.M. Sequential hemifield gating of alpha- and beta-behavioral performance oscillations after microsaccades. J Neurophysiol 118, 2789-2805 (2017).